Bevezetés
- A nemzetközi irodalom áttekintése
- A hazai irodalom áttekintése
- Kísérleti célkitűzés
- A téma jelentősége
- A vizsgálat tárgya
- A hazai pelefajok jellemzése
- Mogyorós pele (Muscardinus avellanarius)
- Nagy pele (Glis glis)
- Erdei pele (Dryomys nitedula)
- A vizsgálati terület
- A terület leírása
- A fás legelők jelentősége
- Anyag és módszer
- A kutatás módszere
- A csapdák ismertetése
- A mérés ismertetése
- A kutatási eredmények
- A 2000-es eredmények
- A 2001-es eredmények
- A 2002-es eredmények
- A 2003-as eredmények
- A kutatás összesítése
- A kutatás értékelése
- Felosztás korcsoportokra
- Hazai standard adatokkal való összehasonlítás
- Összehasonlítás a nemzetközi adatokkal
- Összehasonlítás Andéra (1987) csehországi adataival
- Következtetések, javaslatok
- A vizsgálattal azt szeretném megvizsgálni, hogy a kisgombosi öregtölgyesben mennyire stabil a mogyorós pele és erdei pele populációja,
- A testméreteik mennyire illeszkednek bele a hazai standard méretekbe.
- Szeretném kimutatni, hogy a morfometriai eredmények tükrében, mely európai populációhoz állnak közel az egyes fajok hazai populációi.
- Remélem, hogy a több éves kutatómunkám elősegíti a terület mihamarabbi védettségét, mivel hazánkban elég ritka az ilyen nagyszámú erdei pele populáció.
- Farokhossz: a farok tövétől a farok végéig. A farokvégi szőrt nem mérve, nullázott vonalzóval mérve.
- Talphossz: a bal hátsó lábon mérve, sarokízülettől a leghosszabb ujj végéig. Karom nélkül, tolómérővel mérve. Ha az Achilles-inat enyhén benyomjuk akkor az állat kifeszíti a lábujjait így a teljes talp pontosan és biztonságosan lemérhető.
- Fülhossz: A bal fülön mérve tolómérővel a fülnyílás legalsó pontjától a fül csúcsáig.
- Testtömeg: Vászonzsákba helyezve, rugós erőmérővel (Pesola- val) mérve.
Pelepopulációk (Rodentia, Gliridae) vizsgálata
a kisgombosi öregtölgyesben
TESSEDIK SÁMUEL FŐISKOLA
MEZŐGAZDASÁGI VÍZ- ÉS KÖRNYEZETGAZDÁLKODÁSI
FŐISKOLAI KAR
A témát gondozza: Ökológiai Tanszék
Készítette: Sánta Zsolt Levente, IV. évf. hallgató
[HTML - változat]
Összefoglalás
Irodalomjegyzék
Mellékletek
Bevezetés
Mint már a szakdolgozatom címéből (Pelepopulációk (Rodentia; Gliridae) vizsgálata a kisgombosi öregtölgyesben) is kiderül itt két egymással összefüggő dolgot szeretetnék bemutatni.
Egyik az öregtölgyes, amelynek helyi jelentőségű védelmi felterjesztése a pelepopuláció vizsgálatok következménye. Ezt a Hatvani Környezetvédelmi Egyesület (beleértve engem is) terjesztette be.
A másik az ott folyó immár ötéves folyamatos pelekutatás.
Már több mint tíz éve felfigyeltem a kisgombosi öregtölgyes természeti értékeire és elkezdtem felmérni az állat és növényvilágát. Elkezdtem madarászni a területen, és hat évvel ezelött100 db. A és B típusú madárodút helyeztünk ki. Az első ellenőrzés során, nagy meglepetésünkre nagyon sok Mogyorós pele (Muscardinus avellanarius L.) és Erdei pele (Dryomys nitedula P.) került elő az odúkból. Ekkor merült fel bennem egy pelekutató tábor és kutatás ötlete.
A pelékről sajnos még mindig nagyon keveset tudunk, ezért nagyon fontos a pelepopulációk vizsgálata.
Már négy éve vizsgálom e három faj hazai populációinak egyes morfometriai jellemzőit. Vizsgálataim eredményei, a hazai standard testméretek alakulásának mértékére, valamint egyéb európai populációkhoz való hasonlóságára vagy különbségére vonatkozóan vonok le következtetéseket.
Szerettem volna megvizsgálni, hogy a területen a hazai három pelefaj közül megtalálható-e mindegyik, illetve, hogy mennyire stabil az ott található populáció.
A populációvizsgálatot élvefogó facsapdával végeztük és a helyszínen vettük fel az adatokat, amiből szeretném kimutatni, hogy mennyire illeszkedik vagy tér el a saját adataim a hazai standard adatoktól.
A Bükki Nemzeti Parktól megkaptuk a kutatási engedélyt, amelyet kiállítottak erre az évre is.
1. Irodalmi áttekintés
1.1 A nemzetközi irodalom áttekintése
Az Európában folytatott pelékkel kapcsolatos vizsgálatok eredményeit a kutatók egy háromévente megrendezett szakmai fórumon (International Conference on Dormice) vitathatják meg. E viszonylag szűk kutatási terület résztvevői számára a mindennapos kapcsolattartást, naprakész szakmai információt és vitafórumot a The Dormouse Hollow on-line folyóirat biztosítja (www.glirarium.de/dormouse).
Mind a külföldi, mind a hazai szakirodalomban gyakran faunisztikai kutatásokban, bagolyköpet elemzések eredményeként találkozhatunk peleadatokkal (pl. Vohralík und Sofianidou 1987, Purger és Reider 1998).
A nemzetközi szakirodalomban alapvető munkának tekinthető a Niethammer (1978) által szerkesztett Európa emlőseit részletesen bemutató többkötetes kézikönyv pelékkel foglalkozó, G. Storch (1978) által írt fejezete. Ez a munka részletesen foglalkozik az alfajokkal, azok elterjedésével, a fajok őstörténetével, valamint a pelefajok szaporodásával és populációik szerkezetével.
Storch (1978) táblázataiban a testméretek közül a test-, farok, talphossz és helyenként a testtömeg szerepel. A populációkra illetve alfajokra nem átlagértékeket ad meg, hanem a minimum és maximumértékeket, ezeket a nemekre összevontan
Felten és Storch (1965) Görögország kisemlősfaunáját vizsgálva az erdei pele testméreteit adták meg nemek szerinti bontásban.
A populációdinamikájáról született talán legalaposabb vizsgálat R. Juskaitis (1997) munkája. A litván szerző elsősorban a mogyorós pele populációinak dinamikáját tanulmányozta. Igen nagy számú adatait odútelepek kihelyezésével és rendszeres ellenőrzésével gyűjtötte. Testméret adatokat és tömegadatokat vett fel.
Mindazonáltal fontos megállapításokat tesz közzé a mogyorós pele populációinak szerkezetéről, annak éves ingadozásáról és évek közötti változékonyságáról. Hangsúlyozza, hogy a populáció nagyságát a tavasszal szaporodni képes egyedek számán keresztül a téli mortalitás határozza meg és nagyság akár ötödére-nyolcadára is csökkenhet, több éven keresztül viselve egy-egy kedvezőtlen (enyhe, hómentes) tél következményeit. A szaporodás éves ritmusára vonatkozó adataival összhangban állnak saját hazai megfigyeléseink is.
Juskaitis (1999) részletesen elemzi a mogyorós pele testtömegének éves változásait. Eredményei azt mutatják, hogy az állatok tömege az ébredés után júniusig csökken, ezután állandósul, majd augusztusban kezd erőteljesen növekedni a téli álomra felkészülésként.
A Bright és Morris (1989, 1990) az egyetlen Angliában őshonos pelefaj, a mogyorós pele védelmére és az ehhez szükséges élőhely preferencia vizsgálatokra koncentráltak kutatásaik során. Ők a 15,5 g-nál könnyebb példányokat tekintik fiatalnak (Bright and Morris 1992.).
Filippucci és Civitelli (1988) figyelmeztet arra, hogy a külső morfológiai bélyegek alapján megállapított populációk közötti rokonsági kapcsolatokat nem feltétlenül támasztják alá a genetikai vizsgálatok. Azonos morfológiai jellegzetességek létrejöhetnek különböző kariotípusokból is, azonos vagy hasonló szelekciós nyomás hatására.
Filippucci és Kotsakis (1994) az erdei pele sajátos elterjedési mintázatára hívja fel a figyelmet. Míg a többi európai pelefaj a nyugati Palearktikum jellegzetessége, addig az erdei pele hatalmas elterjedési területe fragmentált, számos teljesen elszigetelt populációt alkot.
Filippucci és Zima (1994) cikkében megemlíti, hogy a kerti pelétől eltekintve az európai pelefajok kariotípusa kis változatosságot mutat. Az erdei pele esetében különösen feltűnő ez a homogenitás, mivel e faj elterjedési területe hatalmas, ugyanakkor erősen fragmentált. Ennél a fajnál az átlagtól lényegesen különböző nagyon kis populációk, illetve egyes egyedek nyomára bukkantak, ezek különbözősége azonban elszigeteltségük miatt genetikai sodródás eredménye lehet. Az erdei és a kerti pele genetikai változatosságát alfajaik számával (19 ill. 10) összevetve a szerző megállapítja, hogy a taxonómiai változatosság nem korrelál a kariotípusok változatosságával. Hozzáteszi ugyanakkor, hogy a szín- és testméret változatossága ezzel szemben korrelál (Zima et al. 1994).
Krystufek és Vohralík (1994) az erdei pele teljes európai elterjedését feldolgozó, az addigi publikált adatokat összefoglaló munkájában Magyarország gyakorlatilag fehér folt a térképen.
1.2 A hazai irodalom áttekintése
A Gliridae családdal foglalkozó hazai szakirodalom sokkal szegényesebb. Magyarországon, nagyon kevés pelefajokkal foglalkozó tanulmány jelent meg.
Vásárhelyi (1934) saját megfigyelései alapján részletes leírást ad a mogyorós pele életmódjára vonatkozóan. Bár stílusa és színvonala napjaink kutatóinak talán már nem elfogadható, ritkasága miatt mégis értékes publikációról van szó. Ebben leírja, hogy megfigyelései szerint a mogyorós pele április elején ébred téli álmából, fiatal (nem feltétlenül önálló!) állatokat az év során május, július, augusztus és szeptember hónapokban talált. Megjegyzi továbbá, hogy a szeptemberi kölykezés csak “bő bükkmakkos esztendőkben” fordul elő.
Számos faunisztikai adatot találunk bagolyköpet vizsgálatok eredményei között, melyek nagyban segítettek a fajok magyarországi elterjedési térképének megrajzolásában (Bakó 1996). Ilyenek Andrési (1983), Kalotás (1986, 1989), Haraszty (1984), Purger (1997, 1998) bagolyköpet adatai.
Egyéb faunisztikai kutatásokban alkalmanként szintén találkozhatunk peleadatokkal (Pl. Vásárhelyi 1932, 1959, Czajlik 1988). Ezeket az adatokat a pelékre összpontosítva először Schmidt (1974) kísérelte meg összegezni.
Czajik (1988) hang alapján állapította meg a nagy pele jelenlétét a Mátra nyugati részén és ezt a módszert populációdinamikai vizsgálatokra is alkalmasnak tartja.
Az első átfogó jellegű magyarországi pelefajokkal foglalkozó munka Bakó (1996) munkája. Az MTM Emlőstárának katalógusadatait és saját csapdázási eredményeit felhasználva megállapította a pelefajok fontosabb testméreteinek és koponyaparamétereinek hazai standard értékeit és azok variabilitását (variációs koefficiensét). Eredményei szerint mogyorós pele (79 pld.) esetében a tömeg és a farokhossz, a nagy pelénél (50 pld.) a farok- és fülhossz, az erdei pelénél (16 pld) pedig a test- és talphossz mutatja a legjelentősebb variabilitást.
Gál (1999) szakdolgozatában részletesen elemzi a hazai pelefajok élőhely-preferencia viszonyait. Leírja az egyes fajok által preferált élőhelyek vegetáció-összetételét és struktúráját, valamint ezek alapján javaslatokat dolgoz ki az élőhelyek, így a fajok védelmére.
2. Kísérleti célkitűzés
2.1. A téma jelentősége
Magyarországon mindhárom pelefaj jelentős természetvédelmi értéket képvisel. Mindhárom faj szerepel a Vörös Könyvben, és 1974 óta törvényi oltalom alatt áll (Bankovics és Nechay, 1989). Európai szintű védelmük a Berni Egyezmény III. függeléke alapján biztosított (Bern Convention, 1994).
A pelefajok hazai jelentőségét növeli az a tény, hogy az 1997-ben beindított Nemzeti Biodiverzitás-monitorozó Rendszerben a biodiverzitás értékelésének alapját képző emlősfajok között mindhárom faj szerepel (Horváth, Bakó 1999). Az Nemzeti Biodiverzitás-monitorozó Rendszer kidolgozásával és alkalmazásával hazánk a Riói Egyezményben vállalt kötelezettségét, az élővilág sokféleségének megőrzését és elemeinek fenntartható hasznosítását biztosító nemzeti stratégia és törvénykezés kidolgozását teljesíti. E kötelezettségek teljesítéséhez fel kell mérni az élővilág jelenlegi állapotát, és folyamatosan nyomon követni annak változásait. Ennek során biológusok és természetvédelmi szakemberek vállalták, hogy feltárják az élővilág sokfélesége csökkenésének okait, a Magyar Tudományos Akadémia pedig kidolgozta a biodiverzitás megőrzésének stratégiáját. Különös hangsúlyt kap a biodiverzitás, a fajok sokfélesége és a fajokon belüli variabilitás folyamatos országos nyomon követése, amely kiemelt élőlénycsoportok rendszeres monitorozásán alapul.
A NBmR pelefajok monitorozását kitűző alprojektjében megfogalmazott feladatok között szerepel a pelefajok populációméretének, -sűrűségének és ezek változásának vizsgálata.
Mindezek ellenére egészen napjainkig a magyar zoológiában Csak kevesen végeztek specifikusan a pelékkel kapcsolatos átfogó kutatást (pl. Bakó 1999), az eddigi, pelékre vonatkozó publikált adatok más kutatások, elsősorban bagolyköpet-analízisek “melléktermékei” voltak, vagy nagyon hézagos ismereteket nyújtanak a pelékről. Nagyon keveset tudtunk e kisemlősök morfológiájáról, ökológiai igényéről, elterjedéséről. Bakó (1996) megadja a pelefajok standard koponyaméreteit és testméreteit a Természettudományi Múzeum gyűjteményében található példányok alapján. 1995-ben a Gödöllői Agrártudományi Egyetem Állattani és Ökológiai Tanszékén megkezdődött egy kutatássorozat, melynek célja a magyarországi pelefajok elterjedésének, élőhely-preferenciájának és populációbiológiájának vizsgálata.
2.2. A vizsgálat tárgya
Magyarország területén három pelefaj (Gliridae) fordul elő. Ezek hazai előfordulási gyakoriságuk sorrendjében a mogyorós pele (Muscardinus avellanarius), a nagy pele (Glis glis ) és az erdei pele (Dryomys nitedula ).(Bakó szerint 1996)
2.3 A hazai pelefajok jellemzése
Bár a kutatásaim során csak mogyorós pelét és erdei pelét sikerült eddig kimutatnunk, azért szeretném mind a három pelefajt egy kicsit jellemezni. Nem tartom kizártnak, hogy a területen a nagy pele is előforduljon, mert az élőhelyjelleg megfelene számára is. (Tölgyesekben található meg a leggyakrabban.)
Vagy populációi nem jutottak el a területre
2.3.1 Mogyorós pele (Muscardinus Avellanarius Linnaeus 1766) (1. kép.)

A legelterjedtebb hazai pelefaj (Bakó 1996).
Európa nagyobb részén, így hazánkban is a M. a. avellanarius alfajt találjuk (Storch 1978).
Elterjedési területe Dánia, az Ibériai-félsziget, Észak-Anglia kivételével egész Európát magában foglalja. Északon Svédország déli területeiig, Keleten a Volga középső folyásáig terjed (Storch 1978).
Magyarországon az Északi-középhegységben, a Dunántúli-középhegységben, a Pilisben, a Visegrádi-hegységben, a Vasi- és Zalai-dombságban, Belső-Somogyban, a Somogyi-dombságban, a Dráva mellékén, az Ormánságban, a Baranyai-dombságban, a Mecsekben, a Villányi-hegységben, a Völgység és a Hegyhát erdőségeiben találkozhatunk mogyorós pelével (Bakó 1996).
A szaporodási időszak június elejétől szeptember végéig tart. A magasabban fekvő területeken évente egyszer, az alacsonyabban fekvő területeken évente kétszer szaporodik (Storch 1978). Csehországban áprilistól szeptemberig figyeltek meg szexuálisan aktív hímeket (Kratochvíl 1973). Míg a hímek általában áprilisban ébrednek a téli álomból, addig a nőstények csaknem egy hónappal később, május első felében (Juskaitis 1999). Az Alpok lábánál az első ellés június vége-július elején, a második július-augusztus fordulóján zajlik (Wachtendorf 1951). Litvániai megfigyelések szerint a fiatalok fele május-júniusban, a többi augusztus-szeptemberben születik, és míg áprilistől júniusig nem találkozhatunk független fiatal állatokkal, addig augusztus-szeptemberben a populáció jelentős hányadát ők teszik ki (Juskaitis 1994). Ez a hányad elérheti a 68%-ot (Likhachev 1966). A kölykök 22-24 napos vemhesség után születnek és 30 napos koruk körül hagyják el a fészket. A korábbi ellésekben több, akár hét kölyök születik (Storch 1978).
A populációk mérete jelentős ingadozást mutat, a populáció sűrűsége akár 8-szorosára csökkenhet vagy növekedhet a téli mortalitástól, illetve a nyári időjárástól függően (Juskaitis 1994). Különösen a hómentes, enyhe teleken magas a téli halálozási ráta, ugyanis a talajba ásott üregekben telelő példányok nem tudják kompenzálni az olvadás és fagyás váltakozását (Likhachev 1966, Juskaitis 1994).
Valódi téli álma októbertől áprilisig tart (Storch 1978).
2.3.2 Nagy pele (Glis glis Linnaeus1766) (2. kép.)

Az emberi közelséget leginkább toleráló, így a leggyakrabban észlelt magyarországi pelefaj. Gyakran találkozhatunk vele épületek környékén, padlásokon, ahol alkalmanként fészket is készít vagy téli álmot alszik (Bakó 1996).
A 8 leírt alfaj közül földrajzi akadályok miatt 3 alfajjal lehetséges a hazai populációk rokonsága. Storch (1978) összefoglaló munkájában megemlíti, hogy a Kelet-Európai populációkra a kisebb testméret jellemző, ezt azonban bulgáriai adatokra alapozva teszi. Elmondható, hogy a hazai populációk rokonságának vizsgálata sem genetikai, sem morfometriai adatok alapján nem történt meg.
A faj elterjedése Európában az Ibériai félsziget déli része, a Benelux államok és Dánia kivételével folyamatos. Angliában egy betelepített populáció él (Thompson 1953, Morris 1997). Északon Riga környékéig, délen Kréta szigetéig, keleten a Pontusz, Kaukázus, Elbrusz hegységekig, illetve Türkmenisztánig húzódik elterjedési területe (Storch 1978).
Magyarországon az Északi-középhegységben, a Dunántúli - középhegységben, a Mecsekben és a Soproni Hegységben találtak nagy pelét (Bakó 1996, MTM Emlőstár katalógusadatok).
Az állatok az első téli álmukat követő tavasszal válnak ivaréretté. A téli álom befejezését követő 4 hét múlva hímek mindegyikében megkezdődik a spermatogenezis (Storch 1978). Hrabe (1970) szlovákiai vizsgálatai szerint a hímek júliusban állnak szexuális aktivitásuk csúcsán. Észak-Morvaországban végzett felmérés szerint júniusban a hímek 69%-a, júliusban 100%-a, szeptemberben 0%-a volt szexuálisan aktív (Gaisler et al. 1977). A 4-6 kölyök 30-32 nap vemhesség után jön világra és további 30 nap után hagyja el a fészket (Storch 1978). Gaisler és munkatársainak (1977) adatai szerint szeptemberben a populáció 23%-át alkotják az egyévesnél fiatalabb állatok. Jóval hosszabb életűek, mint a másik két faj, 6 évig is elélhetnek (Koenig 1960).
Megjegyzendő azonban, hogy a párzás, így az ellés időpontját a téli álom befejezésének ideje határozza meg, így az időjárástól függően évről évre, a populáción belül is változhat. Hideg vagy táplálékszegény nyarakon a teljes szaporodási ciklus kimaradhat (Storch 1978, Bieber 1997). Szeptember vége-október elejétől május-júniusig alszanak téli álmot béleletlen föld alatti üregekben vagy elhagyott odvakban. A téli álom kezdetének időpontja és sikeresen áttelelő példányok aránya az őszi makkterméstől függ. Bő termés esetén később kezdődik a téli álom (Storch 1978). Különösen a fiatal állatok esetében növeli meg a téli mortalitást a gyenge makktermés, mivel nem tudnak elegendő zsírt felhalmozni szervezetükben (Schulze 1970).
2.3.3 Erdei pele (Dryomys nitedula Pallas, 1779) (3. kép.)

Az erdei pele legritkább hazai Gliridae faj (Bakó 1996).
Az irodalom által megkülönböztett alfajok közül hazánkban Storch szerint a D. n. intermedius Nehring 1902 él (Storch 1978). Az alfajok elkülönítése morfometriai bélyegeken alapul. A Kelet-Európai populációk többségének azonban nem ismertek a kariotípusai (Kratochvíl 1973), így biztosan nem jelenthető ki, hogy a Magyarország környezetében előforduló 3 alfaj közül melyikkel állnak közelebbi rokonságban a magyarországi populációk. Mindazonáltal a faj viszonylag fiatal, holocénkori bevándorló Európában (Storch1978, Jánossy 1964), így elképzelhető, hogy a populációk genetikailag még nem különültek el markánsan.
A faj európai elterjedési területe az Alpok nyugati vonulataitól a Volgáig folyamatos. Ázsiában Anatólia és Izrael területétől keleten a Tien-San-, Sinkiang- és Altáj-hegységekig, illetve Pakisztánig találkozhatunk erdei pelével. (Storch 1978, Filippucci 1995). Bár a faj hatalmas területen fordul elő, elterjedése fragmentált. (Filippucci 1995). Ugyancsak a szubpopulációk izolációját írja le Nowakowski és Boratynsky (1999).
Magyarországon előfordulását a Pilisből, a Cserhátból, a Börzsönyből, a Mátrából és a Gödöllői-dombságból jelezték (Bakó 1996).
Az erdei pele második életévétől ivarérett. A szaporodási időszak május-júniusra tehető, ekkor az ivarérett hímek 75%-a mutat szexuális aktivitást. (Gaisler et al. 1977). A párzás májusban zajlik, a kölykök június második felében születnek (Angermann 1963). Bár melegebb éghajlaton (pl. Izraelben) a faj évente 2-3-szor is szaporodhat (Nevo and Amir, 1964.), Magyarországra tapasztalataink szerint inkább az északibb elterjedésű (Lengyelország) populációkhoz hasonlóan az évi egy szaporodási ciklus jellemző (Angermann 1963).
Az almonként 3-5 kölyök egy hónapos korába hagyja el a növényi anyagokból készített fészket, de akár őszig az anyjával maradhat (Storch 1978). Gaisler adatai szerint szeptemberben a populáció 28%-át alkotják a fiatal állatok, melyeket kisebb testtömegük és kevésbé bozontos farkuk alapján lehet elkülöníteni (Gaisler et al. 1977).
Az erdei pele októbertől áprilisig alszik valódi téli álmot a maga készítette fészekben (Storch 1978).
Ugyan egyetlen, macskabagoly köpetből származó adatunk van a kerti pele (Eliomys quercinus Linnaeus,1766) előfordulásáról is 1975-ből az Aggteleki Nemzeti Park területéről (Kordos, 1975), ezt azóta nem erősítették meg, ezért a továbbiakban az első három fajt tekintem hazai pelefajoknak
3. A vizsgálati terület
A kisgombosi öregtölgyes (1. ábra.) Hatvan és Lőrinci város között található. Hatvantól 7 kilométerre északra, Lőrincitől 3 kilométerre délre.
1. ábra. Öregtölgyes térkép
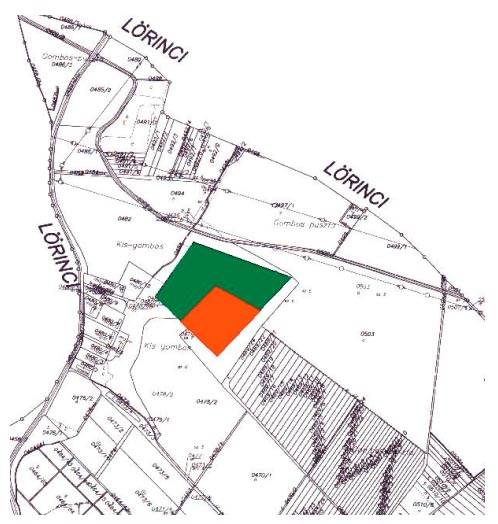
Pirossal jelölve a standard vizsgálati terület
3.1.A terület leírása
Az "öregtölgyes" nevét a területen lévő idős, kb. 200 éves kocsányos tölgyekről kapta.). A mezőgazdasági művelés számára a vízmosásos, meredek oldalakkal rendelkező terület értéktelen, ezért maradhatott máig fenn. A Zagyva-folyó bal partján a Hatvantól Lőrinciig húzódó homok hordalékkúp-síkság utolsó dombja, ezen túl a Délnyugati-Mátraalja vulkanikus képződményei kezdődnek. Természeti képe magán viseli az emberi beavatkozás jeleit: az egykori Zagyva-ártéri keményfás ligeterdőkből felhúzódott tölgyek egymástól 20-30 méter távolságra álló hagyásfák, közöttük a gyepterület dominál, a cserjeszint itt ritkás; a terület egykori fás legelő ma kezeletlen maradványa. Ez a védendő rész magterülete. Ezen kívül nagy kiterjedésben sűrű, vegyes, többnyire vadon élő cserjefajok: kökény, vadrózsa, csíkos kecskerágó, cseregalagonya, keskenylevelű ezüstfa) bozótjai vagy sűrű aljnövényzetű akácligetek találhatók
 Az öregtölgyes a ligetes magterülettel és a hozzá kapcsolódó bozótosokkal kitűnő madárélőhely, amely mind a vonulási, mind a fészkelési időszakban jelentős, de a téli periódusban is jó táplálkozó terület. Mivel mind a nyílt területek, mind a kefesűrűségű bozót és a különböző magassági szintek megtalálhatók, az idős fák némelyike pedig odvas; ragadozó-, magevő és rovarevő énekesmadarak nagy fajszámban tartanak itt revirt. Ehhez hasonló élőhely a hatvani kistérségben nincs.
Az öregtölgyes a ligetes magterülettel és a hozzá kapcsolódó bozótosokkal kitűnő madárélőhely, amely mind a vonulási, mind a fészkelési időszakban jelentős, de a téli periódusban is jó táplálkozó terület. Mivel mind a nyílt területek, mind a kefesűrűségű bozót és a különböző magassági szintek megtalálhatók, az idős fák némelyike pedig odvas; ragadozó-, magevő és rovarevő énekesmadarak nagy fajszámban tartanak itt revirt. Ehhez hasonló élőhely a hatvani kistérségben nincs.
1994 - 2001 között mintegy ötven madárfajt sikerült megfigyelni, melyek rendszeresen előfordulnak a területen. Ez a vegetációtípus kiválóan alkalmas a hazánkban előforduló három pelefajoknak. Ezek közül kettőt (mogyorós pele (Muscardinus avellanarius) és erdei pele (Dryomys nitedula)) a csapdázásaink során sikerült kimutatni. Figyelemre méltó a nagy és stabil erdei pele populáció, amely hazánk legritkább pelefaja. Az öregtölgyes magterületének értékét védett növények előfordulása is emeli: a tavaszi hérics (Adonis vernalis), bunkós hagyma (Allium sphaerocephalon) és a bugás hagyma (Allium paniculatum) található meg. A környékbeli, nagyobb kiterjedésű bozótosok szegélyterületein is előfordulnak védett növények: a budai imola (Centaurea sadleriana), a macskahere (Phlomis tuberosa) és az árlevelű len (Linum tenuifolium).
Élőhelyjellege a Natura Közösség és a Hatvani Környezetvédő Egyesület a terület tulajdonosával (Tangazdaság Rt. Gödöllő) együttműködési megállapodást kötött, melyben a tulajdonos vállalta, hogy a magterületet mai állapotában fenntartja, azon gazdálkodást nem folytat, illetve a fás legelői részen a területjelleg megőrzése érdekében lehetőségei mértékének megfelelően kaszálást, legeltetést folytat, illetve ebbe beleegyezik. Egy évvel ezelőtt a területhez közeli telephellyel rendelkező vállalkozó is megkereste a tangazdaságot azzal, hogy a legelői részen legeltetést végezzen. A tangazdaság ebbe beleegyezett.
Fentiek mellett megfelelő kezeléssel egy jellemző hazai táj: fás legelő fenntartható. A fás legelők száma az országban csekély, ezért a Vörös Könyv Magyarország növénytársulásairól és az Irányelvek a füves területek természetvédelmi szempontú kezeléséhez c. szakmai anyagok védendőként értékelik.
3.2. A fás legelők jelentősége
Az évszázados gazdálkodás a tájat formálva a mai természetvédelem szempontjából értékes élőhelyeket is létrehozott. Ilyen természetátalakító tevékenységnek köszönhetik létüket a fás legelők is.
A fás legelők (legelőerdők) jelentőségét csak az utóbbi években kezdjük felfedezni. Fontosságukat növeli az a tény, hogy a speciális, gyep- és legelőgazdálkodás speciális életközösségeket hozott létre, mely szinte csak az európai kontinensre jellemző. Az emberi tájátalakító munka és a természeti erők összefonódásaként tekinthetünk a legelőerdőkre. Évszázadok viszonylatában is megőrizték arculatukat, mind természetvédelmi, mind tájképi értékekben gazdag területek. Jelenkorunk új természetvédelmi előirányzatainak, mint a biológiai sokféleség megőrzése, mint a fenntartható fejlődés, megfelelnek. Specialitásukból adódóan jelentős felelősség is tevődik a jövő nemzedék vállára: a legelőerdőket csakis további legeltető, kaszáló mezőgazdasági tevékenységgel lehet hosszú távon "ősi" állapotukban fenntartani, kultúrtörténeti és természeti értékeiket megőrizni.
A fás legelők életközösségükben különleges átmenetet képeznek a fátlan gyepek és a zárt erdőtársulások között. Ennek megfelelően igen változatos és nagy fajgazdagságú élővilág alakult ki ezeken a területeken.
A gombosi fás legelőn botanikai szempontból két fontosabb növénytársulás-típus található: a gyepterületek legnagyobb tömegét Thymo-Festucetum tölti ki, kisebb foltokban Campanulo-Stipetum található. Nagy fajgazdagságukat a felmérési eredmények jól mutatják. A felmérések által feltárt természeti érték-leltár is hozzájárulhat a védelemhez
4. Anyag és módszer
4.1 A kutatás módszere
A kutatás élvefogó facsapdázással történt.
A kutatás területe, azaz a csapdák kihelyezése a tölgyes egész területének kb. a negyedén folyt. Ezt a területrészt találtam a legalkalmasabbnak a kutatásra. 2000- ben még csak 50 db csapdánk volt és négy éjszakán át mértünk ezért ennyivel nem tudtuk lefedni az egész területet. 2001-2002-ig már 100 db csapával rendelkeztünk. 2001-ben öt éjszakán keresztül mértünk, míg 2002-ben négy éjszakát. 2003-ban 150 db csapdával négy éjszakán át mértünk.
Ezekben az években nem növeltük a lefedett terület nagyságát, hanem ugyanazon a területen sűrűbben helyeztük el a csapdákat. Ez egy standard területté vált.
4.2 A csapdák ismertetése
 A hagyományos fából készült élvefogó, csapóajtós csapdákat használtunk, melynek minden oldala zárt. Üvegből vágott csapóajtaját az állat akkor hozza működésbe, ha a csapószerkezet belső végére szúrt csalit megmozdítja.
A hagyományos fából készült élvefogó, csapóajtós csapdákat használtunk, melynek minden oldala zárt. Üvegből vágott csapóajtaját az állat akkor hozza működésbe, ha a csapószerkezet belső végére szúrt csalit megmozdítja.
A csapdákat 1-3 méter magasan bokrokon vagy fákon helyeztük el.
Csalinak szezonális gyümölcsöt aprítottunk fel, melyet néha szörppel illetve gyümölcsaromákkal ízesítettünk.
A csapdákat minden nap újracsaliztuk.
4.3 A mérés ismertetése
 A csapdákat naponta kétszer ellenőriztük, éjfélkor és hajnali ötkor. Az állatokat egy jól szellőző vászonzsákba ráztuk ki, ahonnan kézbe véve a következő adatokat mértük le:
A csapdákat naponta kétszer ellenőriztük, éjfélkor és hajnali ötkor. Az állatokat egy jól szellőző vászonzsákba ráztuk ki, ahonnan kézbe véve a következő adatokat mértük le:
Az adatok felvétele előtt mindegyik állat kapott egy egyedi jelölést, melyet kettes számrendszerben a hátán a szőrébe nyírtunk. Így elkerülhetővé vált, hogy ugyanazt az állatot többször is megmérjük. Sajnos ezzel a jelöléssel csak ideiglenes jelölést lehetett alkalmazni, mert ez egy pár hét múlva már eltűnt. Állandó jelölés sajnos még nem működik tökéletesen. Voltak próbálkozások madárgyűrűvel a lábon, de azt nagyon hamar lerágták. Napjainkban kutatások folynak a fültetoválás bevezetése terén.
 Az összes adatot a helyszínen vettünk fel, minden példányról a helyszínen kitöltöttünk egy általunk összeállított adatlapot.), amely az alábbi adatokat tartalmazza: dátum, helyszín, időpont, csapda száma, az állat faja, neme és méretei, időjárás, valamint a csapda helye (növényfaj, magassága) és a körülötte található vegetáció.
Az összes adatot a helyszínen vettünk fel, minden példányról a helyszínen kitöltöttünk egy általunk összeállított adatlapot.), amely az alábbi adatokat tartalmazza: dátum, helyszín, időpont, csapda száma, az állat faja, neme és méretei, időjárás, valamint a csapda helye (növényfaj, magassága) és a körülötte található vegetáció.
5. A kutatási eredmények
5.1 A 2000-es eredmények
2000-ben még csak 50 darab csapdával fogtunk bele a kutatásba és nem vártunk ekkora mennyiségű fogásra. Érdekes, hogy ekkor még a mogyorós pele volt (15 példány, +1 visszafogás) a több (1.-2. táblázat.) mint az erdei pele (9 példány, +6 visszafogás) (3.-4. táblázat.)
1. táblázat. Mogyorós pelefogás 2000-ben
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
A csapda nővénye |
A csapda magassága (m) |
|
07.04 |
Esti |
♂ |
1. |
63 |
13,2 |
16,2 |
Tölgy |
1,5 |
|
07.05 |
Esti |
♀ |
2. |
59 |
11,9 |
17,6 |
Galagonya |
1,0 |
|
07.05 |
Esti |
♀ |
3. |
65 |
11,6 |
15,6 |
Galagonya |
1,5 |
|
07.05 |
Esti |
♀ |
4. |
54 |
11,5 |
16,1 |
Tölgy |
2,0 |
|
07.05 |
Esti |
♂ |
5. |
64 |
11,3 |
15,8 |
Gyepűrózsa |
1,3 |
|
07.05 |
Esti |
♂ |
6. |
67 |
11,2 |
14,0 |
Galagonya |
1,5 |
|
07.07 |
Reggeli |
♀ |
7. |
64 |
11,7 |
15,7 |
Tölgy |
2,0 |
|
07.07 |
Reggeli |
♂ |
8. |
Letört |
11,4 |
15,9 |
Galagonya |
1,7 |
|
07.07 |
Reggeli |
♂ |
9. |
64 |
11,8 |
16,7 |
Galagonya |
1,5 |
|
07.07 |
Esti |
♀ |
10. |
63 |
12,2 |
15,8 |
Galagonya |
1,3 |
|
07.07 |
Esti |
♀ |
11. |
65 |
11,9 |
15,9 |
Kökény |
1,0 |
|
07.08 |
Esti |
♂ |
12. |
64 |
12,1 |
15,9 |
Tölgy |
1,8 |
|
07.08 |
Esti |
♀ |
13. |
Letört |
13,0 |
15,9 |
Galagonya |
1,0 |
|
07.08 |
Esti |
♂ |
14. |
63 |
11,2 |
15,9 |
Galagonya |
1,5 |
|
07.08 |
Esti |
♂ |
15. |
71 |
12,2 |
18,3 |
Galagonya |
1,0 |
2. táblázat. Mogyorós pele visszafogás 2000-ben
|
Dátum |
Időpont |
Nem |
Jel |
A csapda növénye |
A csapda magassága(m) |
|
07.08 |
Reggeli |
♂ |
9. |
Galagonya |
1,8 |
3. táblázat.
Erdei pelefogás 2000-ben|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
A csapda nővénye |
A csapda magassága (m) |
|
07.06 |
Reggeli |
♀ |
1. |
89 |
13,0 |
22,2 |
Vackor |
1,3 |
|
07.06 |
Reggeli |
♀ |
2. |
69 |
9,8 |
15,0 |
Galagonya |
1,5 |
|
07.06 |
Reggeli |
♂ |
3. |
71 |
10,0 |
18,6 |
Galagonya |
1,8 |
|
07.07 |
Reggeli |
♀ |
4. |
71 |
12,7 |
21,4 |
Galagonya |
1,5 |
|
07.07 |
Esti |
♂ |
5. |
82 |
12,1 |
21,5 |
Galagonya |
1,0 |
|
07.07 |
Esti |
♂ |
6. |
91 |
13,2 |
22,3 |
Tölgy |
1,7 |
|
07.07 |
Esti |
♀ |
7. |
82 |
12,1 |
21,1 |
Galagonya |
1,5 |
|
07.08 |
Esti |
♀ |
8. |
83 |
11,6 |
21,6 |
Kökény |
1,2 |
|
07.08 |
Esti |
♂ |
9. |
91 |
13,2 |
23,3 |
Tölgy |
2,0 |
4. táblázat.
Erdei pele visszafogás 2000-ben|
Dátum |
Időpont |
Nem |
Jel |
A csapda növénye |
A csapda magassága(m) |
|
07.07 |
Reggeli |
♂ |
3. |
Galagonya |
1,5 |
|
07.07 |
Esti |
♀ |
4. |
Galagonya |
1,7 |
|
07.08 |
Esti |
♂ |
3. |
Galagonya |
1,0 |
|
07.08 |
Esti |
♀ |
4. |
Galagonya |
1,5 |
|
07.09 |
Reggeli |
♀ |
2. |
Galagonya |
2,0 |
|
07.09 |
Reggeli |
♀ |
7. |
Tölgy |
1,0 |
5.2 A 2001-es eredmények
2001-ben 100 darab csapdával is alig fogtunk mogyorós pelét (2 példány, +1 visszafogás) (5.-6. táblázat.). Viszont nagy örömünkre a ritka erdei peléből viszont annál többet (16 példány, +21 visszafogás) (7.-8. táblázat.)
5. táblázat. Mogyorós pelefogás 2001-ben
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
A csapda nővénye |
A csapda magassága (m) |
|
07.08 |
Reggeli |
♀ |
1. |
67 |
11,3 |
16,1 |
Galagonya |
1.8 |
|
07.09 |
Reggeli |
♂ |
2. |
66 |
10,9 |
19,9 |
Galagonya |
2,0 |
6. táblázat.
Mogyoróspele visszafogás 2001-ben|
Dátum |
Időpont |
Nem |
Jel |
A csapda növénye |
A csapda magassága(m) |
|
08.01 |
Reggeli |
♂ |
2. |
Kökény |
1,5 |
7. táblázat. Erdei pelefogás 2001-ben
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
A csapda nővénye |
A csapda magassága (m) |
|
07.28 |
Reggeli |
♀ |
1. |
66 |
11,8 |
22,2 |
Galagonya |
2,0 |
|
07.28 |
Reggeli |
♂ |
2. |
82 |
10,5 |
21,75 |
Galagonya |
1,8 |
|
07.28 |
Reggeli |
♀ |
3. |
74 |
10,1 |
22,05 |
Tölgy |
1,5 |
|
07.29 |
Reggeli |
♀ |
4. |
74 |
12,4 |
19,95 |
Galagonya |
1,0 |
|
07.29 |
Reggeli |
♀ |
5. |
82 |
15,0 |
23,0 |
Galagonya |
1,3 |
|
07.29 |
Reggeli |
♂ |
6. |
81 |
14,0 |
23,3 |
Galagonya |
1,2 |
|
07.29 |
Esti |
♀ |
7. |
88 |
12,9 |
22,55 |
Galagonya |
2,2 |
|
07.29 |
Esti |
♂ |
8. |
86 |
15,5 |
22,8 |
Galagonya |
2,0 |
|
07.29 |
Esti |
♀ |
9. |
86 |
14,45 |
22,95 |
Tölgy |
2,0 |
|
07.29 |
Esti |
♀ |
10. |
76 |
12,0 |
21,85 |
Tölgy |
1,8 |
|
07.30 |
Esti |
♀ |
11. |
79 |
13,2 |
21,35 |
Tölgy |
1,0 |
|
07.30 |
Esti |
♂ |
12. |
91 |
13,25 |
22,45 |
Galagonya |
1,2 |
|
07.30 |
Esti |
♂ |
13. |
78 |
14,35 |
22,5 |
Galagonya |
1,0 |
|
07.30 |
Esti |
♂ |
14. |
79 |
13,4 |
22,9 |
Galagonya |
1,2 |
|
07.31 |
Esti |
♂ |
15. |
74 |
12,8 |
22,7 |
Galagonya |
1,5 |
|
07.31 |
Esti |
♀ |
16. |
85 |
13,2 |
21,6 |
Galagonya |
1,3 |
8. táblázat. Erdei pele visszafogás 2001-ben
|
Dátum |
Időpont |
Nem |
Jel |
A csapda növénye |
A csapda magassága(m) |
|
07.29 |
Esti |
♀ |
5. |
Galagonya |
1,2 |
|
07.29 |
Esti |
♀ |
4. |
Kökény |
1,0 |
|
07.29 |
Esti |
♀ |
1. |
Galagonya |
1,8 |
|
07. 30 |
Reggeli |
♀ |
3. |
Tölgy |
2,0 |
|
07. 30 |
Reggeli |
♀ |
4. |
Galagonya |
1,5 |
|
07. 30 |
Reggeli |
♀ |
1. |
Galagonya |
2,2 |
|
07. 30 |
Esti |
♂ |
8. |
Galagonya |
1,2 |
|
07.30 |
Esti |
♀ |
5. |
Galagonya |
1,8 |
|
07.30 |
Esti |
♂ |
6. |
Galagonya |
1,4 |
|
07.30 |
Esti |
♀ |
4. |
Kökény |
1,0 |
|
07.30 |
Reggeli |
♀ |
3. |
Galagonya |
1,5 |
|
07.31 |
Reggeli |
♀ |
4. |
Akác |
1,3 |
|
07.31 |
Reggeli |
♀ |
11. |
Galagonya |
1,5 |
|
07.31 |
Reggeli |
♂ |
12. |
Galagonya |
1,0 |
|
07.31 |
Reggeli |
♀ |
10. |
Galagonya |
1,8 |
|
07.31 |
Reggeli |
♀ |
1. |
Olajfa |
1,5 |
|
07.31 |
Reggeli |
♂ |
8. |
Tölgy |
2,1 |
|
07.31 |
Esti |
♀ |
1. |
Olajfa |
1,5 |
|
07.31 |
Esti |
♀ |
9. |
Galagonya |
1,3 |
|
08.01 |
Reggeli |
♂ |
14. |
Tölgy |
2,0 |
|
08.01 |
Reggeli |
♀ |
1. |
Olajfa |
1,5 |
5.3 A 2002-es eredmények
2002-ben ismét 100 darab csapdával kezdtük a kutatást és szinte ugyanannyi mogyorós pelét (6 példány, +2 visszafogás) (9.-10. táblázat.) sikerült fogni, mint erdei pelét (7 példány, +1 visszafogás) (11.-12. táblázat.)
9. táblázat. Mogyorós pelefogás 2002-ben
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
Súly (g) |
A csapda növénye |
A csapda magassága (m) |
|
07.22 |
Esti |
♂ |
1. |
70 |
14,6 |
17,2 |
45,2 |
Galagonya |
1,8 |
|
07.23 |
Esti |
♂ |
2. |
60 |
12,2 |
16,95 |
18,9 |
Galagonya |
2,0 |
|
07.24 |
Reggeli |
♀ |
3. |
71 |
12,75 |
16,4 |
16,7 |
Tölgy |
2,0 |
|
07.24 |
Reggeli |
♂ |
4. |
71 |
13,2 |
17,45 |
17,1 |
Kökény |
1,0 |
|
07.24 |
Esti |
♂ |
5. |
73 |
13,45 |
15,8 |
16,9 |
Kökény |
1,2 |
|
07.25 |
Reggeli |
♀ |
6. |
69 |
12,5 |
15,9 |
16,5 |
Tölgy |
2,0 |
10. táblázat.
Mogyorós pele visszafogás 2002-ben|
Dátum |
Időpont |
Nem |
Jel |
Súly (g) |
A Csapda növénye |
A csapda magassága (m) |
|
07.24 |
Esti |
♂ |
1. |
17,2 |
Kökény |
1,8 |
|
07.25 |
Reggeli |
♂ |
1. |
15,7 |
Galagonya |
1,2 |
11. táblázat. Erdei pelefogás 2002-ben
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
Súly (g) |
A csapda növénye |
A csapda magassága (m) |
|
07.22 |
Esti |
♂ |
1. |
76 |
14,4 |
22,2 |
25,0 |
Galagonya |
1,5 |
|
07.22 |
Esti |
♂ |
2. |
81 |
15,5 |
21,05 |
24,3 |
Galagonya |
1,2 |
|
07.23 |
Reggeli |
♂ |
3. |
87 |
12,1 |
21,4 |
24,2 |
Galagonya |
2,0 |
|
07.23 |
Reggeli |
♀ |
4. |
84 |
11,9 |
21,0 |
30,0 |
Galagonya |
1,0 |
|
07.23 |
Esti |
♂ |
5. |
81 |
13,4 |
21,3 |
35,0 |
Galagonya |
1,3 |
|
07.23 |
Esti |
♀ |
6. |
75 |
14,7 |
20,85 |
32,4 |
Galagonya |
1,5 |
|
07.24 |
Reggeli |
♂ |
7. |
96 |
12,4 |
21,6 |
33,2 |
Galagonya |
1,2 |
12. táblázat.
Erdei pele visszafogás 2002-ben|
Dátum |
Időpont |
Nem |
Jel |
Súly (g) |
A Csapda növénye |
A csapda magassága (m) |
|
07.25 |
Reggeli |
♂ |
5. |
32,0 |
Kökény |
1,8 |
5.4 A 2003-as eredmények
2003-ban már sikerült 150 darab csapdával kimenni a területre, de az egyedszám nem növekedett arányosan a csapdák számával. Mogyorós peléből fogtuk megint a kevesebbet (6 példány, +1 visszafogás) (13.-14. táblázat.), míg erdei peléből (10 példány, +4 visszafogás) (15.-16. táblázat.)
13. táblázat. Mogyorós pelefogás 2003-ban
|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
Súly (g) |
A csapda növénye |
A csapda magassága (m) |
|
07.23 |
Esti |
♂ |
1. |
71 |
12,2 |
16,7 |
16,7 |
Galagonya |
1,5 |
|
07.23 |
Esti |
♀ |
2. |
73 |
12,85 |
16,1 |
15,2 |
Galagonya |
1,2 |
|
07.24 |
Reggeli |
♀ |
3. |
75 |
12,4 |
15,9 |
19,3 |
Gyepűrózsa |
1,0 |
|
07.24 |
Reggeli |
♂ |
4. |
70 |
12,4 |
16,3 |
15,3 |
Galagonya |
1,0 |
|
07.25 |
Esti |
♂ |
5. |
77 |
12,9 |
17,0 |
15,2 |
Galagonya |
1,3 |
|
07.26 |
Esti |
♂ |
6. |
62 |
9,8 |
14,3 |
13,0 |
Galagonya |
1,5 |
14. táblázat.
Mogyorós pele visszafogás 2003-ban|
Dátum |
Időpont |
Nem |
Jel |
Súly (g) |
A Csapda növénye |
A csapda magassága (m) |
|
07.26 |
Reggeli |
♀ |
3. |
18,3 |
Galagonya |
1,8 |
15. táblázat.
Erdei pelefogás 2003-ban|
Dátum |
Időpont |
Nem |
Jel |
Farok (mm) |
Fül (mm) |
Talp (mm) |
Súly (g) |
A csapda növénye |
A csapda magassága (m) |
|
07.23 |
Esti |
♀ |
1. |
85 |
14,4 |
21,4 |
34,1 |
Galagonya |
1,5 |
|
07.24 |
Esti |
♂ |
2. |
72 |
11,2 |
20,9 |
21,4 |
Tölgy |
2,0 |
|
07.24 |
Esti |
♂ |
3. |
73 |
12,2 |
21,1 |
25,0 |
Galagonya |
2,0 |
|
07.25 |
Esti |
♀ |
4. |
91 |
14,1 |
21,8 |
34,0 |
Galagonya |
1,0 |
|
07.25 |
Esti |
♀ |
5. |
78 |
12,2 |
20,2 |
19,6 |
Galagonya |
1,3 |
|
07.26 |
Reggeli |
♂ |
6. |
82 |
11,2 |
22,2 |
27,8 |
Galagonya |
1,5 |
|
07.26 |
Reggeli |
♀ |
7. |
85 |
11,4 |
21,5 |
22,7 |
Olajfa |
1,5 |
|
07.26 |
Reggeli |
♀ |
8. |
90 |
12,1 |
21,4 |
23,0 |
Galagonya |
1,2 |
|
07.26 |
Esti |
♂ |
9. |
81 |
11,4 |
22,3 |
26,7 |
Tölgy |
2,0 |
|
07.26 |
Esti |
♀ |
10. |
76 |
13,1 |
21,9 |
31,2 |
Kökény |
1,2 |
16. táblázat.
Erdei pele visszafogás 2003-ban|
Dátum |
Időpont |
Nem |
Jel |
Súly (g) |
A Csapda növénye |
A csapda magassága (m) |
|
07.25 |
Reggeli |
♂ |
2. |
20,9 |
Galagonya |
1,8 |
|
07.25 |
Esti |
♂ |
3. |
23,0 |
Galagonya |
1,0 |
|
07.27 |
Reggeli |
♀ |
8. |
25,2 |
Galagonya |
1,5 |
|
07.27 |
Reggeli |
♂ |
3. |
23,7 |
Galagonya |
2,0 |
5.5 A kutatás összesítése
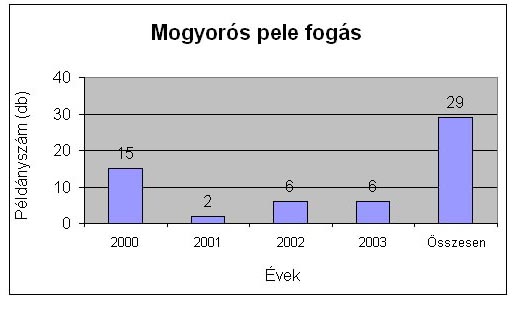
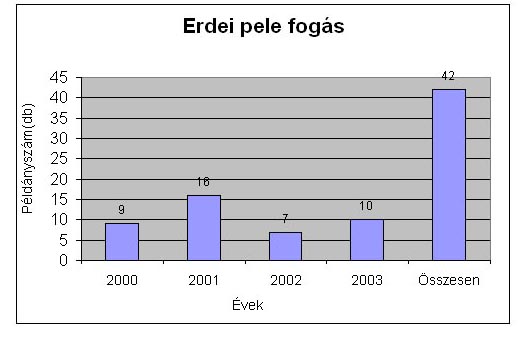
A négy év alatt fogott mogyorós és erdei pele adatokat, egyedszámra és évekre lebontva a 17. táblázat. tartalmazza. Összesen 29 darab mogyorós pelét és +5 darab visszafogást sikerült fogni négy év alatt, míg erdei peléből ugyanannyi idő alatt 42 darabot és +32 visszafogást. Egész Magyarországon elég ritka olyan területet találni, ahol ennyi erdei pele előfordul. Az 3-4. ábrán ábrázolom a mogyoróspele és erdei pele fogásokat.
17. táblázat. Összesen fogott pelék száma
|
Év |
Csapdák száma (db) |
Mogyorós pele (db) |
Erdei pele (db) |
||
|
|
|
Fogás |
Visszafogás |
Fogás |
Visszafogás |
|
2000 |
50 |
15 |
1 |
9 |
6 |
|
2001 |
100 |
2 |
1 |
16 |
21 |
|
2002 |
100 |
6 |
2 |
7 |
1 |
|
2003 |
150 |
6 |
1 |
10 |
4 |
|
Összesen |
|
29 |
5 |
42 |
32 |
3. ábra Mogyorós pele fogások összesítve
4. ábra Erdei pele fogások összesítve
6. A kutatás értékelése
Ebből a 29 példány mogyorós pele adatsorból és 42 példány erdei pele adatsorból azt szeretném kimutatni, hogy az általam mért testméretek, mennyire illeszkednek, illetve térnek el a hazai standard testméretektől, illetve szeretném összehasonlítani a rendelkezésemre álló nemzetközi adatsorokkal is.
6.1 Felosztás korcsoportokra
A korcsoportokra osztás célja az volt, hogy kiküszöböljem a nem kifejlett állatok adatai által okozott értéktorzulást a standard testméretek esetében. Andéra (1986, 1987) megállapította, hogy gyakorlatilag egyetlen testméret sem vizsgálható az állat korától függetlenül. Adatai szerint a növekedés a testtömeg esetében egyes fajoknál eléri a 120%-ot, a farokhossz esetében a 35%-ot, a fül- és talphossznál pedig a 20%-ot. Andéra eredményeit figyelembe véve a standard testméretek számításánál nem tekinthettem el a fiatal egyedek adatainak kizárásától.
A nemzetközi irodalomban a standard méretek megadásánál vagy egyáltalán nem különítenek el korcsoportokat (Storch 1978) vagy testtömeg alapján következtetnek az egyedek korára (Bright and Morris 1992) vagy több mint két korcsoportot különböztetnek meg (Andéra 1986, 1987). Pontos kormeghatározás fogkopás alapján lehetséges (Gaisler et al., cit. Andéra 1986, 1987), erre azonban nekem nem volt lehetőségem, mivel élvefogó csapdát használtam.
A nemzetközi gyakorlatnak (Sidorowicz 1959, Andéra 1986, 1987, Bieber 1997), megfelelően az adott évben született, tehát még át nem telelt állatokat tekintettem fiatalnak és a legalább egy telet túlélt állatokat kifejlettnek. Bár a pelefajok testméretei első életévük után is nőnek (Andéra 1986, 1987, Bright and Morris 1992, Bieber 1997), pusztán statisztikai módszerekkel nem láttam megvalósíthatónak további korcsoportok elkülönítését.
A fiatalok és kifejlett állatok statisztikai elkülönítését megelőzően a fajok szaporodási mintázatának ismeretében kiválogattam a bizonyosan kifejlett példányokat és ezeket én soroltam korcsoportba. A továbbiakban csak a kifejlett korcsoportba tartozó és az előzetesen elkülönített egyedek adatival dolgoztam.
Így az elkülönítés után 24 darab mogyorós pele adatom és 37 darab erdei pele adatom maradt, amit össze kell hasonlítanom a hazai standard adatokkal, illetve a nemzetközi adatokkal. A testtömeg adatsorom azért sokkal hiányosabb, mert 2000-2002-ig nem tudtam testtömeget mérni.
Kicsit kevésnek tűnhet ez az adatsor, viszont ha belegondolunk, hogy a nemzetközi kimutatásnál ennek csak a töredékével dolgoztak akkor már nem is, tűnik olyan kevésnek. Sőt a hazai standard testméretek megadását Bakó-Kosztra (1999) is csak 35 darab erdei peléből állapította meg.
6.2 Hazai standard adatokkal való összehasonlítás
Bakó-Kosztra 1999-ben megadta a hazai pelefajok standard testméreteit. Ezek az adatok saját csapdázásaiból mások csapdázási adataiból és emlőstári adatokból tevődtek össze.
Bakó-Kosztra (1999) mogyorós pele statisztikai adatait a 18. táblázatban foglaltam össze, míg a saját mogyorós pele adataimat a 19. táblázatban közlöm.
18. táblázat. A mogyorós pele hazai standard adatai (Bakó-Kosztra 1999)
|
Farokhossz (mm) |
Fülhossz (mm) |
Talphossz (mm) |
Testtömeg (g) |
|
|
Példányszám (db) |
109 |
121 |
121 |
104 |
|
Maximum |
86,00 |
15,70 |
23,00 |
30,00 |
|
Minimum |
45,50 |
7,25 |
10,52 |
10,00 |
|
Átlag |
65,76 |
11,062 |
16,121 |
18,60 |
|
Szórás |
6,474 |
1,447 |
1,866 |
3,012 |
|
Variancia |
41,916 |
2,093 |
3,483 |
9,073 |
19. táblázat. A mogyorós pele hazai statisztikai adatai (saját adat)
|
|
Farokhossz (mm) |
Fülhossz (mm) |
Talphossz (mm) |
Testtömeg (g) |
|
Példányszám (db) |
24 |
24 |
24 |
11 |
|
Maximum |
77 |
14,6 |
18,3 |
19,3 |
|
Minimum |
60 |
11,2 |
14 |
15,2 |
|
Átlag |
67,88 |
12,28 |
16,23 |
16,83 |
|
Szórás |
4,42 |
0,83 |
0,81 |
1,36 |
|
Variancia |
19,51 |
0,68 |
0,66 |
1,85 |
Az általam mért adatok szinte alig térnek el a hazai standard adatoktól. Az eltérés valószínűleg az eltérő példányszámmal magyarázható.
Bakó-Kosztra (1999) erdei pele hazai statisztikai adatait a 20. táblázatban foglaltam össze, míg a saját erdei pele adataimat a 21. táblázatban közlöm.
20. táblázat. Az erdei pele hazai standard adatai (Bakó-Kosztra 1999)
|
Farokhossz (mm) |
Fülhossz (mm) |
Talphossz (mm) |
Testtömeg (g) |
|
|
Példányszám (db) |
35 |
34 |
35 |
31 |
|
maximum |
95 |
17,5 |
23,5 |
49 |
|
minimum |
62 |
10 |
18,35 |
20 |
|
átlag |
78,57 |
12,24 |
21,02 |
33,73 |
|
szórás |
7,713 |
1,758 |
1,119 |
7,222 |
|
variancia |
59,487 |
3,089 |
1,252 |
52,156 |
21. táblázat. Az erdei pele hazai statisztikai adatai (saját adat)
|
|
Farokhossz (mm) |
Fülhossz (mm) |
Talphossz (mm) |
Testtömeg (g) |
|
Példányszám (db) |
37 |
37 |
37 |
15 |
|
Maximum |
96 |
15,5 |
23,3 |
35 |
|
Minimum |
71 |
10,1 |
19,95 |
19,6 |
|
Átlag |
82,19 |
12,96 |
21,87 |
27,88 |
|
Szórás |
6,14 |
1,30 |
0,79 |
5,00 |
|
Variancia |
37,71 |
1,70 |
0,62 |
25,01 |
Lényeges különbség a testméret adatokban az erdei pelénél sem tapasztalható. A különbségek oka lehet az is, hogy míg Bakó-Kosztra adatai különböző területekről származnak addig az én adataim egy helyről. Ezért lehetséges, hogy testméret Magyarországon belül is változik a különböző populációk között.
A másik különbség oka lehet, hogy én csak nyári adatokkal dolgoztam.
6.3 Összehasonlítás nemzetközi adatokkal
A szakirodalmi forrásokban fellelhető adatok jellegüknél fogva nem teszik lehetővé az összehasonlítások statisztikai alátámasztását. Egyfelől a források többsége csak minimum és maximum értéket közöl, átlagot és elemszámot esetenként, szórásértékeket pedig egyetlen forrásban sem találtam. Másrészt a szerzők nagyon kis minta (n=3-17) alapján állapítják meg egy-egy földrajzi egység populációinak jellemzőit, míg az én mintáim ennél lényegesen nagyobbak (n=24-37), így ha a nemzetközi gyakorlathoz igazodva minimum-maximum értékeket adok meg, lényegesen nagyobb mérettartományt jelölök meg, mint az összehasonlítandó adatok. Az egyetlen általam ismert forrás, amely alaposabb összehasonlítást tesz lehetővé, Andéra (1986, 1987) átfogó tanulmánya, melynek adataival részletesen összevetem eredményeimet a 6.3.1 fejezetben.
A 22.-23. táblázatban összehasonlítom a nemzetközi és hazai adatokat.
22. táblázat. Hazai és nemzetközi adatok összehasonlítása (Mogyorós pele)
|
Származási hely |
Méret |
Farokhossz (mm) |
Talphossz (mm) |
|
Lengyelország M. a. avellanarius |
Min.-max. Átlag |
61-65 63 |
14-17 16 |
|
Észak, Közép és Nyugat Európa |
Min.-max. |
55-82 |
15-17,5 |
|
Németország |
Min.-max. Átlag |
53-75 67 |
15-18 16 |
|
Belgium |
Átlag |
73 |
15 |
|
Közép és Dél Olaszország M. a. speciosus |
Min.-max. |
62-94 |
15-17 |
|
Észak Görögország M. a. zeus |
Min.-max. |
69 |
14-15 |
|
Magyarország (Bakó-Kosztra 1999) |
Min.-max. Átlag |
45,5-86,0 65,76 |
10,52-23,0 16,12 |
|
Magyarország (saját adat) |
Min.-max. Átlag |
60-77 67,88 |
14-18,3 16,23 |
A mogyorós pele hazai populációjának testméret-átlagai szinte tökéletesen megegyeznek a németországi populáció jellemzőivel. Ez alátámasztja Storch (1978) feltételezését, miszerint Magyarországon is a M. a. avellanarius alfaj él.
23. táblázat. Hazai és nemzetközi adatok összehasonlítása (Erdei pele)
|
Származási hely |
Méret |
Farokhossz (mm) |
Talphossz (mm) |
Testtömeg (g) |
Forrás |
|
Olaszország |
Min.-max. |
75-86 |
19-20 |
25-26 |
Storch (1978) |
|
Kelet Alpok |
Min.-max. |
76-90 |
18-21 |
20-28 |
Miller (1912) Storch (1978) |
|
Görögország |
Min.-max. |
62-95 |
17-22 |
17,5-26,4 |
Ondrias (1966) |
|
Bulgária |
Min.-max. |
84-94 |
19-20,5 |
- |
Sidorowicz (1959) |
|
Lengyelország |
Min.-max. |
78-91 |
20-22 |
16,9-22,8 |
Sidorowicz (1959) |
|
Macedónia |
Min.-max. |
78-89 |
19-21 |
- |
Storch (1978) |
|
Montenegro |
Min.-max. |
79-83 |
19-21 |
- |
Storch (1978) |
|
Szrebia |
Átlag |
75 |
17 |
- |
Storch(1978) |
|
Csehország |
Min.-max. Átlag |
78,97 |
21,28 |
17-27 25,80 |
Andéra(1987) |
|
Magyarország |
Min.-max Átlag |
62-95 78,57 |
23,5-35 21,02 |
20-49 33,73 |
Bakó-Kosztra (1999) |
|
Magyarország |
Min.-max. Átlag |
71-96 82,12 |
19,95-23,3 21,87 |
19,6-35 27,88 |
Saját adat (2003) |
Az erdei pele esetében nem állapítható meg egyértelmű hasonlóság egyik európai populációval sem. A csehországi populációhoz hasonlóan a hazai populáció átlagos testtömege lényegesen nagyobb bármely leírt európai populáció testtömeg-átlagánál. Bár csupán tájékozódás szintjén, de megállapítható továbbá, hogy a hazai populáció egyedei európai viszonylatban nagyobb termetűek közé tartoznak.
6.3.1. Összehasonlítás Andéra (1987) csehországi adataival
Eredményeimet összehasonlítottam Andéra (1986, 1987) általam feldolgozott csehországi adataival (24-25. táblázat.). Felhasznált munkájában Andéra mindhárom fajnál négy korcsoportot határozott meg, ezek közül az elsőt (2-3 hónapos, első téli álom előtt) feleltettem meg az általam fiatalnak nevezett csoportnak, a másik hármat (1-4 éves) összevontam. A korcsoportok átlagai és az elemszámok ismeretében kiszámoltam a három csoport átlagát és ezzel hasonlítottam össze saját kifejlett csoportom testméreteit.
24. táblázat. Mogyorós pele testméretének összehasonlítása
|
Testméret |
Átlag |
|
|
|
Andéra nyomán |
Saját adatok |
|
Farokhossz (mm) |
69,05 |
67,88 |
|
Fülhossz (mm) |
13,67 |
12,28 |
|
Talphossz (mm) |
16,46 |
16,23 |
|
Testtömeg (g) |
18,09 |
16.83 |
Mint Andéra adataiból is kiderül a két ország adatai nagyon, hasonlítanak ezért feltételezhető, hagy Magyarországon is Csehországhoz hasonlóan a M. a. avellanarius alfaj él.
Storch (1978) feltételezése szerint is Magyarországon a M. a. avellanarius alfaj él.
25. táblázat. Erdei pele testméretének összehasonlítása
|
Testméret |
Átlag |
|
|
|
Andéra nyomán |
Saját adatok |
|
Farokhossz (mm) |
78,97 |
82.19 |
|
Fülhossz (mm) |
13,04 |
12,96 |
|
Talphossz (mm) |
21,28 |
21,87 |
|
Testtömeg (g) |
25,80 |
27,88 |
Mind a Csehországi, mind a hazai erdei pele adatok lényegesen nagyobbak, mint a többi Európai adat. Főleg a testtömegbeli különbségek a nagyok.
7. Következtetések, javaslatok
A kutatás eredményeként a kitűzött célt sikerült elérni.
Az általam mért adatok alig különböznek, a hazai eddigi adatoktól.
Magyarországon mindhárom pelefaj jelentős természetvédelmi értéket képvisel. Mindhárom faj szerepel a Vörös Könyvben, és 1974 óta törvényi oltalom alatt áll (Bankovics és Nechay, 1989). Európai szintű védelmük a Berni Egyezmény III. függeléke alapján biztosított (Bern Convention, 1994).
A fiatal és kifejlett állatok elkülönítése csak abban az esetben lehetséges statisztikai módszerekkel, ha bizonyosan kifejlett állatokat előzetesen elkülönítjük. Pusztán testtömeg alapján a csoportok közötti nagy átfedés miatt pontos elkülönítés csak az év bizonyos szakában lehetséges. Ehhez mindenképpen szükség van az egyes fajok hazai szaporodási ciklusának ismeretére. A szakirodalmi adatok alapján a szaporodási időszakok hozzávetőlegesen meghatározhatók, ám mivel ezek ideje és üteme nagymértékben függ az éghajlattól, további pontosító vizsgálatokra van szükség.
Valószínűsíthető, hogy Magyarországon Storch (1978) feltételezése szerint a M. a. avellenarius alfaj él. Több kutatásra és adatra lenne szükség a pontos megállapításra.
Az erdei pelénél már nem sikerült egyértelműen kimutatnom nekem sem, hogy Magyarországon melyik alfaj vagy alfajok élhetnek a 19 törzsalak közül. Annyi biztos, hogy az Európai populációkhoz viszonyítva egy nagyobb testtömegű alfaj található meg hazánkban.
Sajnos a nemzetközi adatok, nem teszik lehetővé a pontos statisztikai összehasonlítást. Szükséges lenne egy európai populációk hasonlóságának, ennek alapján rokonságának morfometriai alapú megállapításához egységes adatközlési módszer bevezetése, de mindenképpen nemzetközi együttműködés szükséges.
Sajnos ezeket az állatokat még mindig nagyon kevesen ismerik és kutatják, úgy hazai szinten, mint nemzetközi szinten.
Ahhoz, hogy taxonómiai következtetéseket lehessen levonni, nem elegendő ennyi adat.
További kutatásokra van szükség, úgy hazai szinten, mint nemzetközi szinten és nemzetközi összefogásra is szükség van.
Összefoglalás
Munkám elsődleges célja a három hazai pelefaj (Gliridae) közül a mogyorós pele (Muscardinus avellanarius) és erdei pele (Dryomys nitedula) hazai standard testméreteinek összehasonlítása a saját általam mért adatokkal. Illetve nemzetközi adatsorokkal való összehasonlítás.
A mogyorós pele, a nagy pele és az erdei pele hazánkban jelentős természetvédelmi értéket képvisel, mindhárom faj törvényes oltalom alatt áll. A pelefajok a Nemzeti Biodiverzitás-monitorozó Rendszerben az aktuálisan veszélyeztetett kategóriába kerültek.
Első lépésként el kellett különítenem a fiatal példányokat, mert ezek adatai torzították volna eredményeimet. Adultnak tekintettem a legalább egyszer áttelelt, tehát ivarérett példányokat. Juvenilisnek az adott évben született példányokat tekintettem. A fogkopás alapján ugyan lehetséges a pelék életkorának becslése (Andéra 1986, 1987), azonban élő állatok esetében ez csak a testméretek alapján, statisztikai módszerekkel történhet. Irodalmi adatok és saját tapasztalataim alapján ismerve a téli álomból való ébredés hozzávetőleges idejét, figyelembe véve a vemhesség időtartamát és a születéstől fészekhagyásig eltelt időt, következtettem a fiatalok lehetséges legkorábbi megjelenésének idejére.
A méréseimből kiderült, hogy az általam mért testméretek, beilleszkednek a hazai standard adatokba. A minimális eltérés oka lehet a különböző számú minta és az, hogy én csak nyári adatokkal dolgoztam.
Irodalomjegyzék
Andéra, M. (1987): Dormice (Gliridae) in Czechoslovakia. Part II.: Muscardinus avellanarius, Dryomys nitedula (Rodentia: Mammalia). Folia Mus. Rer. Natur. Bohem. Occid., Plzen, Zoologica 26, 3-78. p.
Angermann, R. (1963): Zur Ökologie und Biologie des Baumschläfers, Dryomys nitedula (Pallas, 1779) in der Waldsteppenzone. Acta Theriol., 7, 333-367. p.
Bakó B. (1996): Adatok a magyarországi pelefajok morfológiájához, elterjedéséhez és ökológiájához. Szakdolgozat. Janus Pannonius Tudományegyetem, 66 p.
Bakó B., Csorba G., Berty L. (1998): Distribution and ecological requirements of dormouse species ocurring in Hungary. Natura Croatica, 7, 1-9. p.
Bakó B., Kosztra B. (1999): Populations dynamics of dormouse species in Naszály region of Hungary. IVth International Conference on Dormice (Rodentia, Gliridae) 13-16 September 1999 Edirne-Turkey. Book of Abstracts, 6 p.
Bandouk, A. C. & Dos Reis, S. F. (1995): Craniometric variation and subspecific differentation in Thrichomys apereoides in northeastern Brazil (Rodentia: Echimyidae). Z. Saugetierekunde, 60, 176-185. p.
Bankovics A., Nechay G. (1989): Emlősök – Mammalia. In: Rakonczay Z. (1989): Vörös Könyv – A Magyarországon kipusztult és veszélyeztetett növény- és állatfajok. Akadémiai Kiadó, Budapest, 360 pp., pp. 56-60.
Bright, P. & Morris, P. (1989): A practical guide to dormouse conservation. Occasional Publication No. 11. The Mammal Society, London.
Bright, P. W. & Morris, P. (1990): Habitat requirements of dormice in relation to woodland management in southwest England. Biological Conservation, 54, 307-326. p.
Bright, P. W. & Morris, P. (1992): Ranging and nesting behaviour of the dormouse Muscardinus avellanarius, in coppice-with-standards woodland. J. Zool. Lond., 226, 589-600. p.
Czajlik P. (1988): A Mátrai Tájvédelmi Körzet nyugati részének emlős faunája. Fol. Hist. Nat. Mus. Matr., 13, 117-122. p.
Felten, H. & Storch, G. (1965): Insektenfresser und Nagetiere aus N-Griechenland und Yugoslavien. (Mammalia: Insectivora und Rodentia). Senck. Biol., 46 (5), 341-367. p.
Filippucci, M. G. & Civitelli, M. V. (1988): Evolutionary genetics and systematics of the garden dormouse, Eliomys Wagner 1840. 1 – Karyotype divergence. Boll. Zool., 55, 35-45. p.
Filippucci, M. G. & Kotsakis, T. (1994): Biochemical systematics and evolution of Myoxidae. Hystrix, (n. s.) 6 (1-2), 77-97. p.
Gaisler, J., Holas, V., Homolka, M. (1977): Ecology and reproduction of Gliridae (Mammalia) in Northern-Moravia. Folia Zool. 26, 213-228. p.
Haraszty L. (1984): Adatok az uhu (Bubo bubo) Magyarországi táplálkozásviszonyainak Ismeretéhez Puszta, 2 (11), 53-59. p.
Jánossy D. (1964): Letztinterglaziale Vertebraten-Fauna aus der Kálmán Lambrech-Höhle (Bükk Gebirge, NO Ungarn). II. Acta Zool. Acad. Sci. Hungar., 10, 139-195. p.
Juskaitis, R. (1994): The structure and dynamics of common dormouse (Muscardinus avellanarius) populations in Lithuania. Hystrix, (n.s.) 6 (1-2), 273-279. p.
Juskaitis, R. (1997): Breeding of the common dormouse (Muscardinus avellanarius L.) in Lithuania. Nat. Croat., Vol. 6, No. 2, 189-197. p.
Juskaitis, R. (1999): Weight changes of the common dormouse (Muscardinus avellanarius L.) during the year in Lithuania. Book of abstracts. IVth International Conference on Dormice, Edirne, 2 p.
Kratochvíl, J. (1967): Der Baumschläfer, Dryomys nitedula und andere Gliridae-Arten in der Tschecholovakei. Eine ökologische Studie. Zool. listy, 16, 99-110. p.
Kratochvíl, J. (1973): Männliche Sexualorgane und System der Gliridae (Rodentia). Acta Sci. Nat. Brno, 7, 1-52. p.
Likhachev, G. N. (1966): Population structure of common dormouse (Muscardinus avellanarius). Bjul. Mosk. Obsc. Isoyt., Biol. 71, 18-29. p.
Mester Zs. (2002) : Kisgombosi tölgyes védetté nyilvánítási javaslat
Miller, G. S. (1912): Catalogue of the Mammals in Western Europe (Europe exclusive of Russia). British Museum (Natural History), London
Nevo, E., Amir, E. (1964): Geographic variation in reproduction and hibernation patterns of the forest dormouse. J. Mammal., 45, 69-87. p.
Nowakowski, W.K., Boratynski, P. (1999): An attempt to estimate the number and density of Dryomys nitedula population in the Bielowieza Forest. Poster. IVth International Conference on Dormice, Edirne, 2 p.
Ondrias, J. C. (1966): The taxonomy and geographical distribution of the rodents of Greece. Saugetierkundliche Mitteilungen, 14, 1-136. p.
Schmidt E. (2000): Kisemlősök
Schulze, W. (1970): Beitrage zum Vorkommen und zur Biologie der Haselmaus (Muscardinus avellanarius) und der Siebenschläfers (Glis glis) im Südharz . Herczynia (N. F.) 7, 355-371. p.
Sidorowicz, J. (1959): Über Morphologie und Biologie der Haselmaus (Muscardinus avellanarius L.) in Polen. Acta Theriol., 3, 75-81. p.
Storch, G. (1978): Familie Gliridae. In: Niethammer, J. & Krapp, F. (1978): Handbuch der Säugetiere Europas. Band 1. Nagetiere 1. AkademischeVerlagsgesellschaft, Wiesbaden, 201-280. p.
Ujhelyi P. (1998): Emlősállatok cd-rom Kossuth kiadó
Vásárhelyi I. (1934): Adatok a mogyorós pele életmódjának ismeretéhez. Állattani Közlemények, 78-83. p.
Vogel, P. & Frey, H. (1995): L‘hibernation du muscardin Muscardinus avellanarius (Gliridae, Rodentia) en nature: nids, fréquence des réveils et temperature corporelle. Bull. Soc. Vaud. Sc. Nat., 83.3, 217-230.p.
Wachtendorf, W. (1951): Beiträge zur Ökologie und Biologie der Haselmaus (Muscardinus avellanarius L.) in Alpenvorland. Zool. Jb. (Syst.), 80, 189-204. p.
Zima, J., Macholán, M., Filippucci, M. G. (1994): Chromosomal variation and systematics of myoxids. Hystrix, (n. s.) 6 (1-2), 63-76. p.
Zimmermann, K. (1953): Die Rodentia Kretas. Zeit. Säugetierk., 17, 21-51. p.
Mellékletek
(A mellékletek az eredeti dolgozatban megtalálhatók)
Konzulens:
Csizmarik Gábor
Főiskolai tanársegéd
Külső konzulens:
Bakó Botond
Környezetvédelmi és Vízügyi Minisztérium
Természetvédelmi Hivatal
Természetmegőrzési Főosztály
Zoológiai tanácsadó
SZARVAS, 2004